|
|
|||
|
|
Forward. The descriptions of Lamprologus (Lepidiolamprologus) kendalli and L. nkambae occurred nearly simultaneously in the late 1970's with type material of L. kendalli (Poll and Stewart 1977) being obtained northwest of Mutondwe Island, Zambia and L. nkambae (Staeck 1978) being described from the Nkamba Bay population at the Northwestern limit of occurrence for this species, restricted to the Southern shores of Lake Tanganyika. L. nkambae, from Nkamba Bay, was the first of these types available to hobbyists in North America during the early 1980's followed shortly by L. kendalli. Neither species have ever been readily available to cichlid hobbyists likely due to the difficulty obtaining wild material occurring most often as solitary individuals at greater depths, their large adult size, and problems pairing adults for breeding. L. kendalli became more frequently available perhaps because all wild populations encompassing the range outside Nkamba Bay to Kala, Tanzania where considered L. kendalli. Hobbyist mystique seemed to develop concerning L. nkambae with many suggesting L. nkambae or "black nkambae" were darker in coloration, more aggressive, and perhaps possessed different morphology than L. kendalli, often resulting in an increased price of L. nkambae in the hobby. Konings's (1998), observations of L. nkambae and L. kendalli suggested little evidence to support a separate pattern color form or morphology, and he regarded L. nkambae as a junior synonym -owing to the earlier description of L. kendalli- of the species L. kendalli, who's range includes the Nkamba Bay population. New scientific information may question this designation.
|
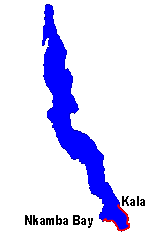
Range of L. nkambae and L. kendalli in Lake Tanganyika. |
|
|
A recent analysis of nuclear and mitochondrial DNA (mtDNA) of Lepidiolamprologus species found L. nkambae specimens were not completely consistent with L. kendalli specimens (Schelly et al. 2005). In the case of L. nkambae and L. kendalli, morphology and nuclear DNA analysis suggest conspecific (same species) arrangement, though mitochondrial DNA (mtDNA) places L. nkambae outside the well defined group of Lepidiolamprologus. The authors interpret this inconsistency in the morphology/nuclear DNA versus mtDNA data as suggesting the Nkamba Bay population of L. nkambae experienced a distant introgressive hybridization with an undetermined Lamprologine species. The genetic signature (haplotype) of this hybridization becoming fixed in the mtDNA of L. nkambae, but via back-crossing and many successive reproductive generations the morphology and nuclear DNA remain similar to L. kendalli. Introgressive hybridization among wild populations of cichlids is not unknown (Salzburger et al. 2002), particularly among introduced populations (Streelman et al. 2004) or where increased turbidity in otherwise transparent waters may prevent visual mate selection among cichlid species who, due to more recent evolution, share few reproductive isolating mechanisms among assemblage members (Seehausen and van Alphen 1998; Seehausen et al. 1997) The authors suggest increased turbidity of Nkamba Bay due to flow from the Lufubu River may have been responsible for introgressive hybridization in the L. nkambae population.
The status of L. nkambae and L. kendalli remains unresolved. The unique genetic character of the Nkamba Bay population suggests a unique and isolated gene pool and designation of a separate species whereas the similar morphology and nuclear material suggest these forms are conspecific. The genetic analysis above was limited to four specimens of the Nkamba Bay population of L. nkambae and a thorough examination of this population and adjacent L. kendalli populations is compelling.
Genetic Analysis Afterword. A few comments on evolution, reproduction, and DNA since most cichlid hobbyists (myself included) may have limited understanding of the genetic analyses performed in the aforementioned study. Organisms possessing Eukaryotic or "complex" cells (members of the kingdoms of animals, plants, fungi, and protista) have genetic material (DNA) found in both the nucleus of the cell and the mitochondria of the cell. Mitochondria are key cell components responsible for the conversion of chemical energy, in glucose for example, into ATP energy which is used for cellular activity, muscle contraction for example. The nuclear DNA directs all cellular activity whereas the DNA in the mitochondria is remnant and thought to be suggestive and the result of a very distant (think billions of years) symbiosis among ancient bacterial cells leading to the evolution of eukaryotic cells. Nuclear DNA is inherited in the gamete cells (egg and sperm) from each parent in animals. Gamete cells in animals undergo cell replication by meiosis during which a unique parceling of genetic material from each of the animals parents occurs via the processes of random assortment and crossing-over along with halving of the chromosome number, rendering each gametic cell produced genetically unique. In contrast, mitochondrial DNA is only inherited from the maternal parent thus each offspring receives mtDNA directly along the maternal line allowing for examination of the code for similarities and difference among specimens/species in the more distant past without the influence of the random parceling of genetic material from both parents during creation of gametic cells which over the short term of several generations tends to make differences in the genetic code more difficult to locate and trace.
|
|||
|
The remainder of this article appeared in Cichlid News 13(2)
Cichlids of the genus Lepidiolamprologus have long drawn my attention due to morphology well-adapted for predation. The cylindrical, elongate body form is particularly reminiscent of the North American Esocids, such as Northern Pike. Lepidiolamprologus kendalli combines this body form, an eye-catching pattern, and a reputation of challenging to maintain in the cichlid aquarium, ensuring continued popularity among hobbyists. Lepidiolamprologus kendalli was once included in the genus Lamprologus. This assemblage of substrate spawning cichlids was recognized to contain many species, some with only distant relationship. Genetic studies revealed the Lake Tanganyika Lamprologines were likely derived from a common riverine ancestor. Taxonomists have more recently restricted the genus Lamprologus to include only riverine forms with the type species Lamprologus congoensis (Sturmbauer et al. 1994). Colombe and Allgayer (1985) published a re-description of the genus Lepidiolamprologus [Pellegrin 1904] and Stiassny (1997) reviewed morphology of Lake Tanganyikan Lamprologines and supported re-designation into the genus Lepidiolamprologus. This genus of nine described species includes larger predatory species (e.g. L. elongatus [genus type species]) and smaller micro-predators (e.g. L. hecqui). Lepidiolamprologus kendalli was described by Poll and Stewart (1977). In the hobby, L. kendalli is also represented by the so-called L. nkambae (“black nkambae”) from Nkamba Bay, Zambia. More recently Konings (1998) reviewed known populations of L. kendalli and found similar variation in color and pattern across all populations and thus little evidence to support L. nkambae as a separate species thus “nkambae” should be treated as a junior synonym of kendalli and identification of this variant as L. kendalli “nkambae”. There are hobbyist reports of differences in morphology and behavior between early imports of so-called L. “nkambae” and L. kendalli yet substantive scientific investigation is lacking, suggesting this issue may remain unresolved until further research is conducted.
L. kendalli is distributed in the southern portion of the Lake Tanganyika along the Tanzanian and Zambian coastlines. Known populations are distributed from Kala, Tanzania around the southern tip of the lake to Nkamba Bay, Zambia. Solitary individuals and pairs have been found inhabiting waters near surface to depths of 45 m, always near the rock substrate and mud interface (Konings 1998).
L. kendalli is a relatively large, monomorphic substrate spawner. Males are generally slightly larger than female broodmates and similar in color and pattern. This species is best housed in at least a four-foot aquarium of 70 gallons or larger. Absolutely essential to this setup is a tight fitting cover. L. kendalli, at a relatively small size (~2"), become highly aggressive towards intraspecific competitors such that an individual may attempt to exclude all other kendalli from its territory by literally driving them out of the aquarium. Even the smallest openings must be covered or plugged; plastic fish bags can accomplish this nicely. This aggressive behavior is fitting with the observations of wild fish only occurring as individuals or pairs. In captivity adults of this species are probably most successfully maintained as lone individuals or in pairs. Raising a group of six or more juveniles will likely ensure both sexes are represented and will allow a pair to form at which point all others may be removed from the aquaria or potentially be eliminated by the dominant pair. Sometimes additional females can be housed with a dominant pair however the behavior of a pair, particularly during spawning, can be unpredictable. Aquarists have reported success with removing a bonded pair to a smaller species only tank however in my experience this inevitably results with loss of the female due to male aggression. A pair may also be maintained in larger community aquaria where spawning and raising offspring can be successful. As a carnivore, this species does well maintained on a high protein, high lipid flake/pellet diet. I have found supplementation of this diet with frozen artemia or mysis essential in conditioning for spawning. L. kendalli seem to prefer a dark enclosed cave for spawning. Clay pots are inexpensive and handy for constructing a spawning cave. This cave may be easily removed for rearing fry, particularly if the pair is housed in a community tank. A spawning cave can be constructed from short (3-4" tall), wide (6" diameter) cylindrical pots. A hole just large enough to accommodate the parent can formed using a drill with masonry bit in the side of the pot near its top (which will become the bottom of the cave). Care should be taken to excavate an opening just large enough to accommodate the largest parent. The small opening size may give the parents a better sense of security and seems preferred over spawning caves with larger openings. On the top of the cave (bottom of the clay pot), another hole may be located (if not already existing on the pot) with a diameter large enough to permit the female, but not the male, to exit. This opening allows the female to escape the male if cornered in the pot. Constructing two nearly identical caves will allow one, along with any brood, to be removed and replaced without greatly disrupting the pair. The eggs of L. kendalli are similar to other Tanganyika substrate spawners and are characteristically small, compared to mouthbrooders, and olive in color. An adult pair may produce 70-150 in a single spawning. Hatching occurs in approximately five days at 78 F and fry are free swimming by 14-16 days. Free swimming fry may be fed a variety of ground flake or pellet feeds. Supplementation with hatched live or frozen baby artemia or any of a variety of dried artemia or artemia replacement products, all of which are high protein, high lipid, seems to greatly enhance survival and support increased growth rates. A variety of suitable tankmates exist for this species, provided a few considerations are made. A four-foot tank is likely the minimum for tank raised specimens whereas wild caught specimens should probably be housed in no less than a six-foot aquarium. Most carnivorous/omnivorous Tanganyika cichlids, large enough not to be consumed, may be considered including: Altolamprologus, Chalinochromis, Julidochromis, Cyprichromis sp. “jumbo” types, and Cyphotilapia, depending upon tank size. Lake Malawi and Lake Victoria haplochromine cichlids also mix well. In my experience, even paired L. kendalli in community aquaria do not defend territory or broods as aggressively as many Neolamprologus species, however wild caught specimens may require more space or fewer tankmates relative to tank raised fish. L. kendalli is available to hobbyists as tank raised juveniles or wild adult fishes. Wild caught fish and tank raised juveniles usually demand a relatively high price despite the fact that adults may produce broods numbering in the 100's. This may reflect the observation of L. kendalli being found at greater depths as lone individuals or pairs, never in great abundance (Konings 1998), and the difficulty of pairing in captivity of both wild and tank raised fish. Whether in pairs or as lone individuals this species makes an interesting, undoubtedly challenging addition to any large cichlid aquarium. Literature Cited Colombe, J., and R. Allagayer. 1985. Description de Variabilichromis, Neolamprologus et Paleolamprologus, generes nouveaux du lac Tanganyika, avec rediscription des generes Lamprologus Schilthuis 1891 et Lepidiolamprologus Pellegrin 1904. Rev. Franc. des Cichlidophiles 49:9_28. Konings, A. 1998. Tanganyika cichlids in their natural habitat, 2nd edition. Cichlid Press, 272pp. Poll M. and D.J. Stewart. 1977. Un nouveau Lamprologus du sud du Lac Tanganika (Zambia) (Pisces, Cichlidae). Revue de Zoologie africaine 91(4): 1047-1056. Salzburger, W., S. Baric, and C. Strumbauer. 2002. Speciation via introgressive hbridization in East African cichlids? Molecular Ecology 11: 619-625. Schelly, R., Salzburger, W., Koblmüller, S., Duftner, N., Sturmbauer, C. (2006). Phylogenetic relationships of the lamprologine cichlid genus Lepidiolamprologus (Teleostei: Perciformes) based on mitochondrial and nuclear sequences, suggesting introgressive hybridization. Molecular Phylogenetics and Evolution, 38(2):426-438. link to PDF Seehausen, O, and JJM van Alphen. 1998. The effect of male coloration on female mate choice in closely related Lake Victoria cichlids. Behaviorial Ecology and Sociology 42: 1-8. Seehausen, O., JJM van Alphen, and F. Witte. 1997. Cichlid diversity threatened by eutrophication that curbs sexual selection. Science 277: 1808-1811. Staeck, W. 1978. Ein neuer Cichlidae aus dem sudlichen Tanganjikasee: Lamprologus nkambae n. sp. (Pisces, Cichlidae). Revue de Zoolie. Africaine 436-441. Stiassny, M.L.J. 1997. A phylogenetic overview of the lamprologine cichlids of Africa (Teleostei,Cichlidae): a morphological perspective. South African Journal of Science, 93: 513-523. Streelman, J. S.L. Gmyrek, M.R. Kidd, C. Kidd, L. Robinson, E. Hert, A.J. Ambali, and T.D. Kocher. 2004. Hybridization and contemporary evolution in an introduced cichlid fish from Lake Malawi National Park. Molecular Ecology 13: 2471-2479. Sturmbauer, C.E., E. Verheyen, and A. Meyer. 1994. Mitochondrial phylogeny of the Lamprologini, the major substrate spawning lineage of cichlid fishes from Lake Tanganyika in Eastern Africa. Molecular Biology and Evolution. 10:751_768.
|
|||
|
Home l Update l Stock List l Gallery | Lake Malawi l Lake Tanganyika l Lake Victoria l Articles l References l Links l About | Contact
© Copyright 1999-2012 All rights reserved.
|
|||